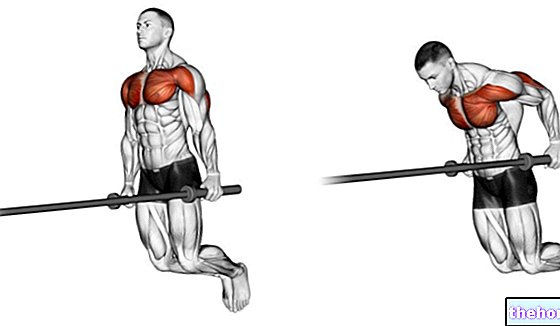
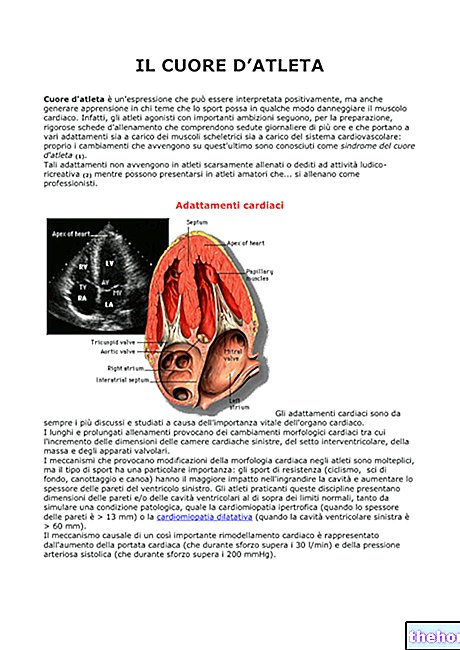
ANMÄRKNINGAR OM EMBRYOLOGI
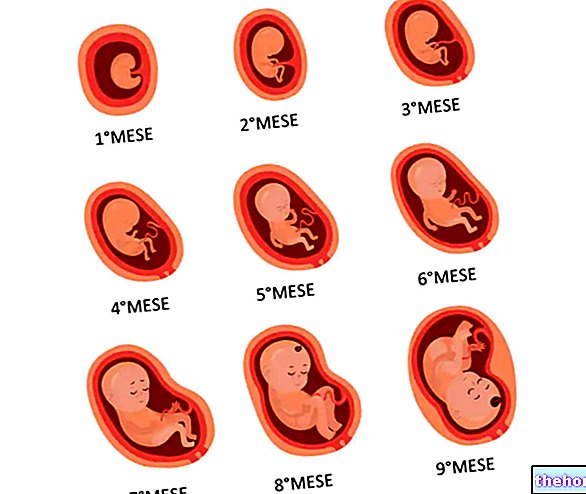
Embryologi studerar sekvensen av utvecklingsformer från zygoten till organismen utrustad med alla dess organ och system.
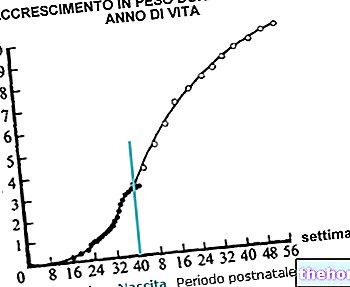
I detta avseende är det värt att komma ihåg skillnaden mellan utveckling (succession av strukturella och organisatoriska faser med ökande komplexitet) och tillväxt, främst avsedd i kvantitativ mening.
I ryggradsdjur metazoans vi bevittnar, stigande i den evolutionära serien upp till människan (genom cyklostomer, fisk, amfibier, reptiler, fåglar och däggdjur), utseendet av vuxna former av ökande komplexitet, för vilken komplikationen av stadierna av embryonal utveckling.
I början delas zygoten, alltid utrustad med reservmaterial, (med efterföljande mitos) i 2, sedan 4, sedan 8, etc. celler som kallas blastomerer, utan tillväxt, tills den når artens normala kärna / cytoplasmatiska förhållande .
Denna inledande segmentering kan följa olika mönster, beroende på mängd och distribution av deutoplasman.
I början är deutoplasman knapp ("oligolecytiska ägg"), så segmenteringen är total och ger upphov till mycket olika blastomerer. När embryonas komplexitet ökar tar det mer tid och material innan dess utveckling tillåter det att börja. självständigt liv. Detta kräver en ökning av deutoplasma ("telolecytiska ägg"), som tenderar att ordnas i en del av zygoten. Detta orsakar en ökande "anisotropi", som är kopplad till modifieringar av segmenteringen, som styrs av två allmänna principer:
- Hertwigs lag säger att vid mitos tenderar den achromatiska spindeln (vars ekvatorn bestämmer delningsplanet för dottercellerna) att vara anordnad i riktning mot den största längden av cytoplasman;
- Balfours lag säger att segmenteringshastigheten är omvänt proportionell mot mängden deutoplasma.
Vi ser då att redan i cyklostomer och i fisk är segmenteringen ojämlik, med en snabbt segmenterad djurpol (som ger embryonets övre strukturer) och en äggula som kommer att innehålla det mesta av reservmaterialet. Denna tendens är ännu större . anisotrop hos amfibier (där det är nödvändigt att förbereda de organ som är ansvariga för luftandning), där äggulestången, medan den långsamt segmenteras, förblir relativt inert och slutligen täcks av celler som härrör från den snabbt segmenterade djurpolen. de viktigaste embryonala stadierna inkluderar: zygote, blastomerer, morula (kluster av blastomerer som liknar en björnbär), blastula (morula med regresserade inre celler), gastrula (blastula där cellerna på ena sidan har invaginerats), där den primitiva kaviteten av organismen, med ett externt cellskikt (ektoderm, från vilket nervsystemet först kommer att härledas) och en int erno (entoderm), mellan vilket ett tredje lager (mesoderm) sedan kommer att placeras. Från dessa lager eller "embryonala ark" kommer sedan, i en ordnad sekvens, att alla vävnader, organ och system.
I ännu mer utvecklade arter är ökningen av deutoplasma (eller "kalv") sådan att den inte ens kan segmenteras. Vi ser alltså att segmenteringen hos fåglar påverkar endast en tunn ytlig skiva, vilket leder till en "discoblastula" och en serie av fenomen som garanterar att embryot bildas på ett annat sätt än det som nämns ovan.
En ytterligare ökning av deutoplasma hade förmodligen inte varit mer effektiv, så hos däggdjur erhålls utvecklingen och tillväxten upp till kapaciteten för självständigt liv med ett annat system.Vi noterar faktiskt hos däggdjur att deutoplasman endast används i de allra första stadierna av utvecklingen; då upprättar embryot metaboliska relationer med moderns organism (med placentans utseende) och använder inte längre deutoplasman, vars överskott elimineras. Vid denna tidpunkt återgår ägget till att bli oligolecitiskt och segmenteringen kan gå tillbaka att vara total (och därför är det i de tidiga stadierna liknande det för "amphioxus), men efter morula fortsätter embryogenesen enligt det mest utvecklade mönstret av fåglar, med en" blastocyst "följt av implantation på livmoderväggen, så att metabolismen av embryot säkras av moderorganismen (via moderkakan) snarare än av deutoplasman.
EMBRYO DIFFERENTIATION
När segmenteringen av zygoten har fört kärnan / cytoplasmatiska förhållandet till artens norm måste tillväxten också börja, parallellt med utvecklingen.Därför börjar metabolismen, med utseendet av nukleoli och proteinsyntes. Den på så sätt initierade proteinsyntesen beror på generna som är ansvariga för de tidiga stadierna av embryonal utveckling. Dessa gener är deprimerade av de ämnen som finns i de olika blastomererna på djur- och kalvpolen. I sin tur kan produkterna från dessa tidiga gener nedpressa operonerna hos generna som ansvarar för de senare stadierna. Produkterna från denna andra serie av gener kommer att kunna verka både i betydelsen att konstruera nya embryonala strukturer och i den meningen att undertrycka de tidigare operonerna och avpressa följande, i en ordnad sekvens som leder till konstruktionen av den nya organismen , tack vare den genetiska information som samlats in. från genomet genom årtusenden till allt mer utvecklade arter.
Haeckels berömda uttryck "" ontogeni sammanfattar fylogeni "uttrycker faktiskt det faktum att de högre arterna upprepar, i stadierna av embryonisk utveckling, den succession som redan finns i de evolutionärt föregående arterna.
De tidiga stadierna av embryot tenderar att vara lika hos ryggradsdjur, särskilt upp till gälarnas utseende.
Hos arter som övergår till luftandning absorberas sedan gälarna och återanvänds (till exempel för bildning av endokrina körtlar), men den genetiska informationen om bildandet av gälarna bevaras också hos människor. Detta är uppenbarligen ett exempel på embryonala strukturgener som finns i genomet hos alla ryggradsdjur och måste förbli undertryckta efter att ha fungerat i sitt ontogenetiska ögonblick.
Tolkningen av embryogenes i betydelsen av reglering av genverkan gör det möjligt att förena de komplexa traditionella erfarenheterna av experimentell embryologi.
TVILLINGAR
Zygoten och de första blastomererna, tills proteinsyntesen börjar, är totipotenta, det vill säga kan ge liv åt en hel organism. Till detta är experimenten från Spemann kopplad, som fick två embryon från strypningen av en amfibisk zygot. Ett liknande fenomen dyker upp på grundval av fenomenet identiska tvillingar hos människan, som just av denna anledning kallas monozygotiska (MZ). Spemanns experimentella tvillingar var halva storleken på det normala, medan de hos människan är helt normala. Detta förklaras eftersom de två embryona måste dela den enda äggulan som redan har mottagits i amfibierna, medan embryon hos mannen kan ta emot allt som är nödvändigt för deras utveckling och tillväxt genom moderkakan.
Det bör komma ihåg att hos "människan har två tredjedelar av fallen med tvillingar" ett annat ursprung: de härrör från en och annan samtidig mognad av två folliklar, med frisättning av två ägglossningar som, när de befruktas, ger två zygoter; faktiskt, i i detta fall talar vi om dizygotiska tvillingar (DZ).
Eftersom MZ -tvillingarna, dividerade med mitos från den enda zygoten, har samma genom måste skillnaderna mellan dem vara av miljömässigt ursprung.I stället är genomet för två DZ -tvillingar bara lika mycket som för två bröder. Tvillingmetod, används ofta inom mänsklig genetik och även inom idrottsområdet.
I "mannen, där vissa etiska skäl skulle förbjuda experimenterande, kan det fastställas hur mycket någon karaktär regleras av ärftliga faktorer: i själva verket är strängt ärvda karaktärer (såsom blodgrupper) alltid överensstämmande endast med tvillingar i MZ; att överensstämmelse för ett drag i MZ är nära det för DZ, det dras att miljöfaktorer råder över ärftliga faktorer för att bestämma den fenotypiska egenskapen.