Virus kan bara replikera inuti en värdcell, utnyttja dess metaboliska apparat och använda sin egen genetiska information; emellertid sker multiplikation endast i celler som är mottagliga för viruset, det vill säga försedda med specifika ytreceptorer och som kan utföra de replikativa faserna av dess genom.
Multiplikationsprocessen är indelad i olika faser:
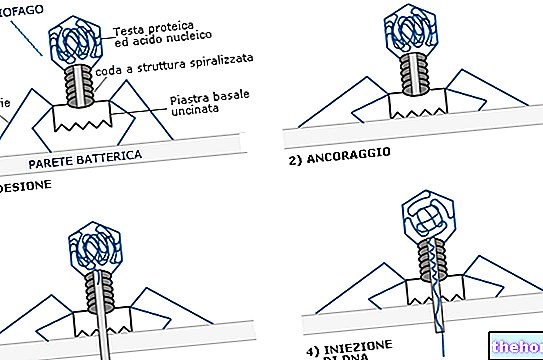
1: a fas: attack eller adsorption av viruset till cellmembranet;
2: a steget: virusets penetration eller dess nukleinsyra i cellens cytoplasma;
3: e fas: avklädning eller förmörkelse (förlust av virushöljen och exponering av nukleinsyra);
4: e fas: replikation (syntes av makromolekyler, dvs DNA, RNA och virala proteiner); virus har olika replikativa strategier och var och en av dem multiplicerar på ett annat sätt, utnyttjar enzymer och organeller i värdcellen;
5: e fasen: sammansättning (bildning inuti cellen - i kärnan eller i cytosolen - av kapsiden; inuti detta hölje sätts det virala DNA in och bildar nukleokapsiden);
Sjätte fasen: frisläppande eller flykt av viruset från cellen.
Med några undantag är den virala replikationscykeln mycket snabb och slutförs på 8-24 timmar. Var och en av dessa faser är komplex och typisk för varje art; i själva verket finns det en stor variation av replikativa strategier och mekanismer; de två första (adsorption och penetration) och den sista (exit), till exempel, är olika beroende på om viruset har en perikapsid eller inte. Medan bakteriofager injicerar sina nukleinsyror direkt i värdcellens cytoplasma, kommer dessa djur in genom pinocytos och frigörs både genom celllys och pinocytos; under denna passage förvärvar de nya virionerna fosfolipidskiktet och efter att de lämnat kan de infektera nya celler.
Virusattack, penetration och replikering
Naken virus kommer in i cellen genom mikropinocytos, även kallad viropepsis, det vill säga med samma biologiska mekanism som den använder för att internalisera korpuskulära ämnen under 1 μm. Väl i cytoplasman smälter cellulära proteaser kapsiden och nukleinsyra (viralt DNA) släpps ut i cytoplasman.
Virusets attack på cellen förmedlas av proteiner som kallas antireceptorer, som finns på den virala kapsiden och på den virala perikapsiden, som känner igen molekyler eller proteiner som finns på cellytan och kallas receptorer. Adsorptionsfasen medieras därför av interaktionen mellan antireceptor och receptor.
ADSORPTION: stereokemisk interaktion mellan specifika kemiska grupper som exponeras på den yttre ytan av den mottagliga cellen (receptorer) och av virionen (antireceptorer).
HIV, till exempel, angriper främst T-hjälparlymfocyter, eftersom det har antireceptorer som känner igen specifika proteiner som exponeras på deras cellyta.Antireceptorn för HIV-viruset är ett perikapsid glykoprotein, kallat GP120, medan det för T-lymfocyten kallas CD- 4 ; av denna anledning är T -hjälparlymfocyten också känd som T4. När viruset väl är bundet kan det komma in i cellen på två sätt:
fusion utifrån: perikapsiden smälter ihop med cellmembranet och släpps ut i cytoplasman (typiskt för HIV och belagda virus);
fusion inifrån: viruset kommer in i en vesikel genom pinocytos. Väl i cytoplasman smälter perikapsiden samman med vesikelmembranet och kapsiden släpps ut i cytoplasman, som till exempel händer med influensaviruset och i allmänhet med nakna.
Som förväntat finns det många variationer i hur viruset kommer in i värdcellen.
REPLIKATION: virus har olika replikativa strategier, betingade av typen av nukleinsyra som finns i kapsiden; under replikation genererar virus i allmänhet två typer av proteiner: tidigt (av enzymatisk och reglerande karaktär, såsom polymeraser) och sent (strukturellt, vilket kommer att bilda kapsid och perikapsid). I vilket fall som helst innebär replikation först och främst en "förändring och omdirigering av värdens metabolism", vilket gör att viruset kan multiplicera sitt eget genom.
Det sista steget är utträdet av de nya virusen från cellen (som följer kapsidens sammansättning i kärnan eller i cytoplasman). I allmänhet kommer nakna virus ut genom celllys; i de belagda, dock vissa virala proteiner under replikering, ansvarig för bildandet av perikapsiden, går för att sätta in sig på ett av värdcellens membran (till exempel det cytoplasmatiska membranet, det nukleära eller Golgimembranet eller det endoplasmatiska retikulumet); på detta sätt, efter vid självmontering närmar sig nukleokapsiden det modifierade membranet, den spirande processen börjar och viruset slipper svepa in sig delvis i det modifierade membranet och förvärva perikapsiden (eller kuvertet).
VIRUS-CELL-INTERAKTIONSMEKANISMER: virusinfektion.
PRODUKTIONSINFEKTION: producerar nya virus (virala avkommor);
RESTRIKTIV: viruset multipliceras endast när cellen befinner sig under vissa förhållanden (till exempel i fas S);
ABORTIV: viruset replikerar inte utan uttrycker bara några proteiner utan att kunna ge upphov till nya virioner;
PERSISTENT: det kan vara kroniskt - viruset replikerar långsamt och cellen släpper ut viruset under långa perioder (månader eller till och med år), som vid HIV och kronisk hepatit - eller latent (virusgenomet förblir tyst i kärnan i värdcell under långa perioder, bara för att återaktiveras för att ge en produktiv infektion, som i fallet med herpes simplex eller zoster).
TRANSFORMERING: typiskt för onkogena virus, som inte dödar cellen utan omvandlar den i neoplastisk mening. I dessa fall integreras det virala genomet i det cellulära och tar namnet provirus; denna förändring kan leda till en genetisk förändring av värdcellen, som transformeras i neoplastisk mening och, genom att sprida sig på ett okontrollerat sätt, överför anomalierna till dottercellerna.
Viral infektion kan orsaka akut sjukdom med kort behandling och okomplicerad återhämtning (vanligtvis på grund av en produktiv infektion, som vid förkylning), eller kronisk sjukdom.
Andra artiklar om "Virusmultiplikation"
- Virusstruktur och klassificering
- Virus
- Sjukdomar orsakade av virus och antivirala läkemedel